Species: 種の分類
ラビリンチュラ類の種の分類についても,やはり問題が多い。基本的に細胞の形態が単純であり,さらに形態の可塑性も大きいため,種の同定基準となる形質 を認識することが難しい。Raghukumar (1988) は,生育環境に近い,海水に松花粉を浮かべたものを,形態観察のための「基準培地」とすることを提唱している。形質が安定して観察され,現れる形態も比較 的多様であるため,基準培地とすることは有効であると思われる。
現時点では,以下のような特徴を種分類の形質にしていることが多い。
* 細胞壁の有無あるいは厚さ
* 遊走細胞のサイズ
* 遊走細胞の形成様式
* 遊走子嚢のサイズ
* 1つの遊走子嚢の中に形成される遊走細胞の数
* 原基体(proliferous body)の有無あるいは数
* 遊走細胞のサイズ
* 遊走細胞の形成様式
* 遊走子嚢のサイズ
* 1つの遊走子嚢の中に形成される遊走細胞の数
* 原基体(proliferous body)の有無あるいは数
参考: 原基体(proliferous body)
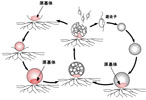 |
Thraustochytrium sp. の生活史。 遊走細胞を放出した後に原基体(proliferous body)を残す種では,このように原基体が発達して新たな栄養細胞を経て,遊走子嚢になることが知られている。 |
ただし,ラビリンチュラ類の 18S rRNA 遺伝子の分子系統樹では,同じ種と同定されている株間の遺伝的距離は,真菌類などに比べても比較的大きく,十分に種が認識できていない可能性もある。
Honda et al. (1998) では,炭素源の資化性について比較して,種を区別するための補助的な特徴とした。しかし,後の分子系統解析結果は,資化性に差のあった株は系統群が異な り,属の階級で区別されることとなった。そのため,資化性の差が分類的にどの階級で有効な形質なのかについては決着がついていない。このような形質が分類 基準となり得るか,という課題から検討する必要があると思われる。
参考: 炭素源資化性
 |
Assimilation of carbon compounds by Aurantiochytrium (= Schizochytrium) limacinum and comparison with several strains of Schizochytrium aggregatum. The values for Aurantiochytrium limacinum are the optical density at 650 nm. From Honda et al. (1998). |